






Introducción:
El género Bacillus comprende bacterias Gram positivas aerobias estrictas o facultativas, formadoras de endosporas resistentes, ubicuas en suelos, sedimentos acuáticos y ambientes vegetales, donde forman parte de comunidades microbianas complejas. Su relevancia agrícola radica en la capacidad de múltiples especies para colonizar la rizósfera y filósfera, secretar metabolitos secundarios con actividad antimicrobiana y modular respuestas defensivas de las plantas hospederas. Kloepper et al., 1978; Sun et al., 2022; Li et al., 2025
En el contexto de biocontrol, bioestimulación y sanidad vegetal, los Bacillus se emplean como alternativas o complementos a los fungicidas químicos, aprovechando mecanismos que incluyen la producción de lipopeptidos (surfactinas, iturinas, fengicinas), enzimas hidrolíticas y elicitores de resistencia inducida. Estos atributos permiten su integración en estrategias de manejo integrado de enfermedades, reduciendo la presión selectiva sobre patógenos y minimizando residuos químicos en cultivos. Sin embargo, la expresión funcional de estos rasgos no es uniforme en el género, lo que justifica un análisis diferenciado por especie y cepa. Sun et al., 2022; Li et al., 2025; Chen et al., 2024
Mecanismos generales de acción de los Bacillus
Mecanismos en el género Bacillus:
La acción de los Bacillus como herramientas biológicas en plantas se organiza en tres ejes principales, ampliamente aceptados en la literatura. Figura 1. Dini-Andreote., 2020; Sun et al., 2022
La acción directa sobre patógenos o el microambiente inmediato de la planta involucra la secreción de metabolitos antimicrobianos y enzimas que interfieren con la viabilidad del inóculo patógeno. Lipopeptidos como surfactinas alteran la permeabilidad de membranas fúngicas, mientras que enzimas hidrolíticas (quitinasas, β-1,3-glucanasas, proteasas) degradan componentes estructurales de paredes celulares. Estos efectos se manifiestan localmente en la rizósfera o filósfera, donde las concentraciones de metabolitos alcanzan umbrales inhibitorios en el espacio compartido por bacterias, patógenos y tejidos vegetales. Li et al., 2025; Sánchez et al., 2023.
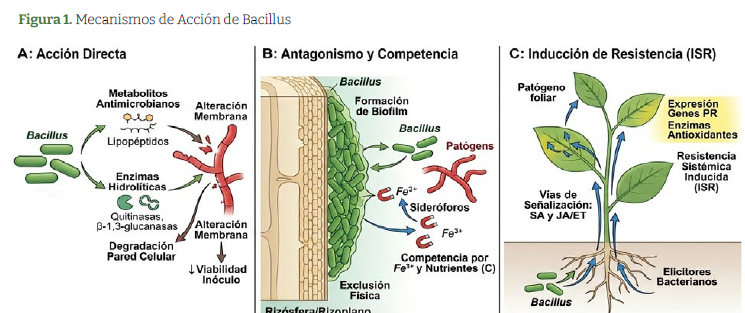
El antagonismo microbiano y la competencia ecológica constituyen un segundo eje, basado en la competencia por sustratos limitantes (hierro, carbono), formación de biofilms que restringen el acceso espacial y producción de sideróforos con alta afinidad por Fe³. La capacidad de Bacillus para formar biofilms densos en superficies radiculares o foliares excluye físicamente a competidores, alterando la composición de la microbiota asociada y reduciendo el espacio nicho disponible a patógenos oportunistas. Sun et al., 2022; Li et al., 2025
La activación de resistencia en la planta hospedera representa el tercer mecanismo, caracterizado por la inducción de respuestas locales o sistémicas mediadas por elicitores bacterianos (lipopéptidos, fragmentos de pared celular, polisacáridos). Estos compuestos activan vías de señalización como la ruta dependiente de salicílico y la ruta jasmonato/ etileno, resultando en la expresión de genes defensivos (PR-genes, enzimas antioxidantes) y mayor resistencia a patógenos en tejidos distales. La distinción entre mecanismos locales (respuestas en el sitio de aplicación) y sistémicos (ISR) es clave, aunque en práctica agronómica los efectos suelen integrarse. Dini-Andreote., 2020
Estos mecanismos son multifactoriales y su contribución relativa depende de la especie bacteriana, la cepa específica, la formulación aplicada, la dosis, las condiciones ambientales y la fisiología del cultivo, lo que explica variaciones entre productos y en las eficacias observadas en campo. Chen et al., 2024; Li et al., 2025
Diferencias entre Bacillus pumilus y otras especies de Bacillus de uso agrícola
Particularidades de la especie Bacillus pumilus
Pasando del género Bacillus en general a la especie Bacillus pumilus, se observan rasgos metabólicos y ecológicos que la distinguen de otras especies agrícolas, aunque con variabilidad significativa entre sus cepas. Wang et al., 2023; Abdurahman et al., 2023; Ma et al., 2023
Características específicas de B. pumilus
No todos los Bacillus producen el mismo perfil de lipopéptidos, enzimas o elicitores, ni exhiben idéntica capacidad de colonización o persistencia ambiental. En B. pumilus, se reporta consistentemente la producción de aminoazúcares, surfactinas, pumilacidinas, bacilisin y enzimas como binasa y quitinasas, que contribuyen a la antibiosis contra fitopatógenos fúngicos como royas, mildeos polvosos y vellosos entre otros. Los genomas de esta especie suelen contener menos clusters biosintéticos NRPS/PKS que en B. velezensis o B. amyloliquefaciens, sugiriendo una estrategia antibiótica más focalizada, aunque igualmente efectiva en ciertos patosistemas. Abdurahman et al., 2023; Ma et al., 2023; García et al., 2025; Wang et al., 2023
En cuanto a colonización, Bacillus pumilus es una bacteria que se establece con facilidad en las raíces, en la superficie de las hojas y dentro de los tejidos de la planta, gracias a que produce esporas, permanece viable en condiciones difíciles y se reactiva cuando el ambiente mejora. En comparación con Bacillus subtilis, suele formar una capa microbiana (biofilm) menos robusta alrededor de las raíces, por lo que su fijación en esa zona puede ser algo menor. Aun así, se adapta muy bien a suelos con la vida microbiana alterada por sequía, uso de agroquímicos o manejo intensivo, y favorece el crecimiento de las plantas sin provocar cambios drásticos en el equilibrio del suelo. Ma et al., 2023; Abdurahman et al., 2023
La inducción de resistencia sistémica (ISR) está documentada en varias cepas de B. pumilus, asociada a lipopeptidos y rutas salicílico y jasmonato/etileno. Comparativamente, B. velezensis destaca por repertorios lipopeptídicos amplios y biofilms rizosféricos intensos; B. subtilis por versatilidad en colonización; B. licheniformis por enzimas y bioestimulación; y B. firmus por efectos nematicidas. Similitudes con estas especies incluyen la esporulación y antibiosis básica, pero B. pumilus presenta diferencias en señalización molecular y afinidad ecológica que justifican su selección específica. Dentro de B. pumilus, las cepas difieren en la abundancia de metabolitos y la intensidad de ISR, resaltando la necesidad de evaluaciones cepa-específicas. Berg et al., 2021;Zhang et al., 2024; Olanrewaju et al., 2017;Wang et al., 2023;Abdurahman et al., 2023;Ma et al., 2023
Sonata: mecanismos de acción y particularidades
Evidencia específica para la cepa QST 2808
Del nivel de especie B. pumilus pasemos ahora al nivel de cepa B. pumilus QST 2808, ingrediente activo de Sonata, distinguiendo lo documentado directamente para este aislado de inferencias generales. Xu et al., 2023; Hovinga., 2014
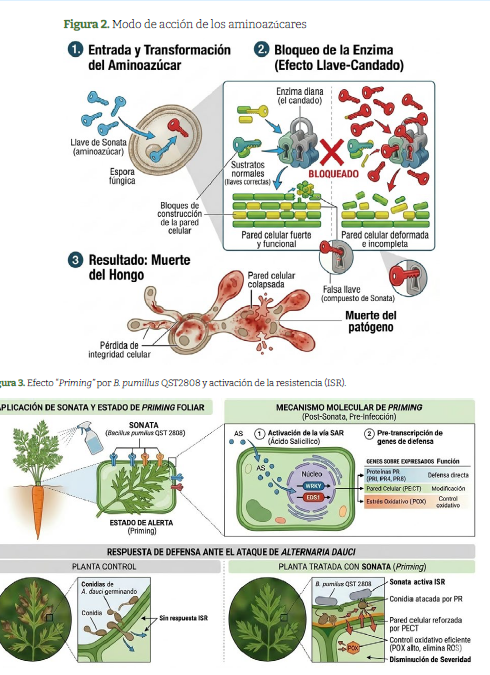
Modo de acción de QST 2808
La cepa QST 2808, aislada de ambientes naturales y producida por fermentación, se aplica como fungicida de contacto para enfermedades foliares. Su modo de acción integra una fase de contacto, donde metabolitos de fermentación inhiben directamente la germinación de esporas patógenas en la superficie foliar, con contribución a la activación de defensas naturales. Evaluaciones regulatorias confirman actividad preventiva y curativa temprana contra royas, mildeos y manchas foliares.Hovinga., 2014; Xu et al., 2023.
Aminoazúcares, activación de resistencia y efecto de cebado
Los aminoazúcares en el producto SonataR cepa QST 2808 se identifican como componentes clave de la actividad fungicida de contacto, alterando estructuras patógenas superficiales. El efecto “llave-candado” de los aminoazúcares sobre las enzimas responsables de la biosíntesis de la pared celular es un claro ejemplo de inhibición enzimática competitiva. Estas enzimas poseen un sitio activo con una conformación tridimensional específica, análoga a una “cerradura”, diseñado para reconocer y unirse a sus sustratos naturales, las “llaves” moleculares correctas. Los aminoazúcares, debido a su similitud estructural con estos sustratos, actúan como llaves falsas que encajan perfectamente en el sitio activo, pero que no pueden ser procesadas. Al ocupar este sitio diana, bloquean físicamente el acceso de los sustratos verdaderos, impidiendo la catálisis y deteniendo la formación de la pared celular, lo que resulta en la pérdida de la integridad celular y, en última instancia, en la muerte del patógeno. Figura 2. Xu et al., 2023; Hovinga., 2014
La activación de respuestas defensivas en plantas tratadas con QST 2808 se evidencia en ensayos funcionales, como en zanahoria frente a Alternaria dauci, donde Sonata reduce severidad de síntomas y activa genes de defensa, clasificándose como Inductor de resistencia (PRI) típico. En genotipos susceptibles, reduce la severidad; en parcialmente resistentes, ofrece protección significativa donde otros biopesticidas fallan. Le Clerc et al.,2025; Pascale et al., 2025; Zhou et al., 2025
El efecto de cebado (priming) describe un estado fisiológico donde la planta, tras exposición a elicitores microbianos, no activa defensas totalmente, pero responde más rápida e intensamente a ataques fungicos posteriores. En zanahoria, plantas tratadas con Sonata muestran menor severidad post-inóculo, con patrones génicos asociados con ISR. Figura 3. Le Clerc et al.,2025.
Discusión
Asumir que todos los productos basados en Bacillus funcionan igual ignora variabilidad en metabolitos, colonización e ISR. La cepa específica determina el perfil: QST 2808 destaca por aminoazúcares en contacto y ISR (efecto priming). La formulación de un producto biológico afecta la estabilidad de los metabolitos y la bacteria; la aplicación de SonataR activa el priming (efecto de cebado o alerta), como evidencian diferentes estudios.Abdurahman et al., 2023; Ma et al., 2023; Pascale et al., 2025; Zhou et al., 2025; Wang et al., 2023; Le Clerc et al.,2025
Bibliografía
Abdurahman, A., et al. (2023). A metagenomic
insight into the rhizosphere microbiota of
healthy and diseased plants. Frontiers in Microbiology,
14. https://doi.org/10.3389/fmicb.
2023.1194606
Berg, G., et al. (2021). The plant microbiome and
native plant species: A review. Frontiers in
Microbiology, 12. https://doi.org/10.3389/fmicb.
2021.8614995
Chen, L., et al. (2024). Synthetic microbial communities:
A new frontier for resilient agriculture.
Frontiers in Plant Science, 15. https://doi.
org/10.3389/fpls.2024.1349357
Dini-Andreote, F. (2020). Endophytes: The role
of extravascular and vascular microorganisms
in plant health. Emerging Topics in Life
Sciences, 4(2), 215–226. https://doi.org/10.1042/
ETLS20200034
García-García, A., et al. (2025). Biopesticides and
their impact on soil microbial diversity. Journal
of Applied Microbiology, 136(3). https://doi.
org/10.1093/jambio/lxae035
Hovinga, S. (2014). Characterization of the Microbiome
associated with the Rhizosphere of
Strawberry (Tesis de Maestría). California State
University, Sacramento. pp. 1-65.
Kloepper, J. W., & Schroth, M. N. (1978). Plant growth-
promoting rhizobacteria on radishes. Proceedings
of the 4th International Conference on
Plant Pathogenic Bacteria, 2, 879-882.
Le Clerc., et al. (2025). Identification of plant resistance
inducers and evaluation of genotype
receptivity for carrot protection against
Alternaria leaf blight. Front. Plant Sci., 04
March 2025. Sec. Plant Pathogen Interactions
Volume 16 – 2025. https://doi.org/10.3389/
fpls.2025.1513301
Li, P., et al. (2025). Integrative omics reveals the
core microbiome of sustainable agroecosystems.
Science of The Total Environment, 905. https://
doi.org/10.1016/j.scitotenv.2025.12913297
Liu, H., Li, J., & Fu, Q. (2023). Rhizosphere microbiome
assembly and its impact on plant fitness.
Frontiers in Plant Science, 14. https://doi.
org/10.3389/fpls.2023.1301431
Ma, Y., et al. (2023). The role of Bacillus species in
biocontrol of plant diseases. Plants, 12(15). https://
doi.org/10.3390/plants12152841
Olanrewaju, O. S., Glick, B. R., & Babalola, O. O.
(2017). Mechanisms of action of plant growth-
promoting bacteria. World Journal of Microbiology
and Biotechnology, 33(11), 197. https://
doi.org/10.1007/s11274-017-2364-9
Pascale, A., et al. (2025). The role of the plant microbiome
in differential disease resistance.
Frontiers in Plant Science, 16, 1513301. https://
doi.org/10.3389/fpls.2025.1513301
Sánchez-López, A. S., et al. (2023). Microbial inoculants
for sustainable agriculture: Current
status and future directions. Microbial Biotechnology,
16(11), 2056-2072. https://doi.
org/10.1111/1751-7915.14348
Sun, X., et al. (2022). Fungal communities in the
rhizosphere of plants under different fertilization
regimes. Microbiology Spectrum, 10(6).
https://doi.org/10.1128/spectrum.02341-22
University of Hertfordshire. (2024). Bacillus
amyloliquefaciens strain QST 713. Pesticide
Properties DataBase (PPDB). http://sitem.herts.
ac.uk/aeru/bpdb/Reports/1701.htm
Wang, J., et al. (2023). Deciphering the rhizosphere
microbiome for disease suppressiveness.
International Journal of Molecular Sciences,
24(16). https://doi.org/10.3390/ijms241612752
Xu, L., et al. (2023). Interaction between plant immunity
and the microbiome. Current Opinion
in Plant Biology, 75. https://doi.org/10.1016/j.
pbi.2023.102432
Zhang, Y., et al. (2024). Advances in endophyte-mediated
plant disease resistance. PeerJ, 12. https://
doi.org/10.7717/peerj.18920
Zhou, J., et al. (2025). Soil microbiome manipulation
for crop health: Mechanisms and applications.
Nature Communications, 16.
Elaborado por: Diego Torres -Kam Flores
